| Evolutsion biologiyaga oid turkumning bir qismi |
| Evolutsion biologiya |
|---|
 |
|
|
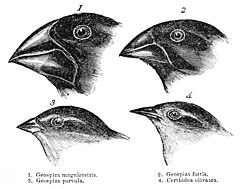
Genetik o‘zgaruvchanlik (genetic variation) — bu individlar (individuals) orasidagi DNK farqi yoki bir turdagi populyatsiyalar (populations) orasidagi farqlardir. Genetik o‘zgaruvchanlikning ko'plab manbalari mavjud bo'lib, ularga mutatsiya va genetik rekombinatsiya kiradi. Mutatsiyalar genetik o‘zgaruvchanlikning asosiy manbai hisoblanadi, biroq genetik dreyf kabi boshqa mexanizmlar ham bunga hissa qo'shadi
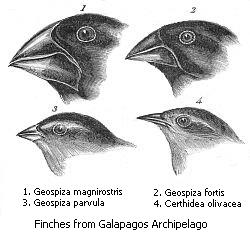
Populyatsiya ichidagi individlar orasida
Genetik o‘zgaruvchanlikni ko'p darajalarda aniqlash mumkin. Genetik o‘zgaruvchanlikni miqdoriy belgilar (ko'plab genlar tomonidan kodlanadigan va uzluksiz o'zgarib turadigan belgilar, masalan, itlarning oyoq uzunligi) yoki diskret belgilar (bir yoki bir nechta genlar bilan kodlanadigan va aniq toifalarga bo'linadigan belgilar, masalan, ba'zi gullarning oq, pushti yoki qizil gulbarg rangi) kabi fenotipik o'zgaruvchanlik kuzatuvlaridan aniqlash imkoniyati mavjud.
Genetik o‘zgaruvchanlikni oqsil elektroforezi (protein electrophoresis) jarayoni yordamida fermentlar darajasidagi o'zgaruvchanlikni tekshirish orqali ham aniqlash mumkin. Polimorf genlar (polymorphic genes) har bir lokusda (locus) bittadan ortiq allelga (allele) ega bo'ladi. Hasharotlar va o'simliklardagi fermentlarni kodlovchi genlarning yarmi polimorf bo'lishi mumkin, umurtqali hayvonlar orasida esa polimorfizm kamroq uchraydi.
Pirovardida, genetik o‘zgaruvchanlik genlardagi nukleotidlarning asoslari ketma-ketligidagi farqlardan kelib chiqadi. Zamonaviy texnologiyalar olimlarga DNK ketma-ketligini bevosita aniqlash imkonini beradi, bu esa ilgari oqsil elektroforezi orqali aniqlanganidan ko'ra ko'proq genetik o‘zgaruvchanlikni aniqlashga yordam berdi. DNKni tekshirish genlarning kodlovchi sohalarida ham, kodlamaydigan intron sohalarida ham genetik o‘zgaruvchanlik mavjudligini ko'rsatdi.
Agar DNK ketma-ketligidagi nukleotidlar tartibining o'zgarishi ushbu DNK ketma-ketligi tomonidan kodlangan oqsillardagi amino kislotalar tartibining farqlanishiga olib kelsa va natijada amino kislotalar ketma-ketligidagi farqlar fermentning shakliga va funktsiyasiga ta'sir qilsa, genetik o‘zgaruvchanlik fenotipik o‘zgaruvchanlikka olib keladi.
Populyatsiyalar orasida
Geografik ajralish natijasida populyatsiyalar o'rtasida yuzaga keladigan farqlar geografik o‘zgaruvchanlik (geographic variation) deb ataladi. Tabiiy tanlanish, genetik dreyf va genlar oqimi geografik o‘zgaruvchanlikka hissa qo'shishi mumkin.
O'lchash
Populyatsiya ichidagi genetik o‘zgaruvchanlik odatda polimorf gen lokuslari foizi yoki geterozigotali individlardagi gen lokuslari foizi sifatida o'lchanadi. Natijalar populyatsiyadagi har bir individning atrof-muhitga moslashish (adaption) jarayonini tushunishda juda foydali bo'lishi mumkin.
Shakllari
Genetik o‘zgaruvchanlik genetik o'zgarishlar asosida yotgan genomik variatsiyaning o'lchami va turiga qarab turli shakllarga bo'linadi. Kichik miqyosdagi ketma-ketlik variatsiyasi (1 kilobazadan kam, kb) asoslar juftining almashinuvi va indellarni (indels) o'z ichiga oladi. Katta miqyosdagi strukturaviy o‘zgaruvchanlik (>1 kb) nusxalar sonining variatsiyasi (yo'qolish yoki orttirish) yoki xromosoma qayta qurilishi (translokatsiya, inversiya yoki segmental orttirilgan uniparental dizomiya) bo'lishi mumkin Ko'chib yuruvchi elementlar va endogen retroviruslar tomonidan amalga oshiriladigan genetik o‘zgaruvchanlik va rekombinatsiya ba'zan xo'jayin genomlarida genetik yangiliklarni keltirib chiqaradigan turli xil turg'un viruslar va ularning defektivlari bilan to'ldiriladi. Butun xromosomalar yoki genomlardagi son jihatidan o'zgarishlar poliploidiya yoki aneuploidiya bo'lishi mumkin.
Populyatsiyalarda saqlanishi
Bir qator omillar populyatsiyalarda genetik o‘zgaruvchanlikni saqlab turadi. Potentsial zararli retsessiv allellar diploid organizmlar populyatsiyasidagi geterozigotali individlarda tanlanishdan yashirin bo'lishi mumkin (retsessiv allellar faqat kamroq uchraydigan gomozigotali individlarda namoyon bo'ladi). Tabiiy tanlanish, shuningdek, muvozanatlashgan polimorfizmlarda (balanced polymorphisms) genetik o‘zgaruvchanlikni saqlab qolishi mumkin. Muvozanatlashgan polimorfizmlar geterozigotalar afzal ko'rilganda yoki tanlanish chastotaga bog'liq bo'lganda yuzaga kelishi mumkin.
RNK viruslari
Korrektura (proofreading) mexanizmining yo'qligi sababli yuzaga keladigan yuqori mutatsiya darajasi RNK viruslari evolyutsiyasiga hissa qo'shadigan genetik o‘zgaruvchanlikning asosiy manbai bo'lib ko'rinadi.. Genetik rekombinatsiya ham RNK viruslari evolyutsiyasi asosida yotgan genetik o‘zgaruvchanlikni hosil qilishda muhim rol o'ynashi isbotlangan Ko'plab RNK viruslari bitta xo'jayin hujayrasida kamida ikkita virusli genom mavjud bo'lganda genetik rekombinatsiyaga kirishishga qodir.. RNK rekombinatsiyasi Picornaviridae ((+)ssRNK) (masalan, poliovirus) guruhida genom arxitekturasini va virusli evolyutsiya yo'nalishini belgilovchi asosiy harakatlantiruvchi kuch bo'lib ko'rinadi. Retroviridae ((+)ssRNK) (masalan, HIV) guruhida RNK genomidagi shikastlanishlar teskari transkriptsiya jarayonida zanjir almashinuvi (genetik rekombinatsiya shakli) orqali chetlab o'tiladi. Rekombinatsiya, shuningdek, Coronaviridae ((+)ssRNK) (masalan, SARS) guruhida ham sodir bo'ladi. RNK viruslaridagi rekombinatsiya genom shikastlanishini bartaraf etishga moslashish (adaptatsiya) bo'lib xizmat qiladi Rekombinatsiya bir turdagi, ammo turli tarmoqlarga mansub hayvon viruslari o'rtasida kamdan-kam hollarda sodir bo'lishi mumkin. Olingan rekombinant viruslar ba'zan odamlarda kasallik tarqalishiga sabab bo'lishi mumkin
Genetik o‘zgaruvchanlik tarixi
Evolyutsion biologlar ko'pincha genetik o‘zgaruvchanlik bilan shug'ullanadilar, bu atama zamonaviy davrda individlar o'rtasidagi DNK ketma-ketligidagi farqlarni anglatadi. Biroq, genetik o‘zgaruvchanlikni miqdoriy baholash va tushunish birinchi to'liq genom ketma-ketligi aniqlanishidan ancha oldin va hatto DNK irsiyat uchun mas'ul molekula ekanligi kashf etilishidan oldin ham yerdagi rang-barang hayotni tushunishga qiziquvchilarning asosiy maqsadi bo'lgan.
Bugungi genetik o‘zgaruvchanlik ta'rifi zamonaviy molekulyar genetikaga tayansa-da, irsiy o‘zgaruvchanlik g'oyasi hatto Charlz Darvinning asarlaridan oldin ham hayotning mazmuni va rivojlanishiga qiziquvchilar uchun markaziy ahamiyatga ega bo'lgan. Irsiy o‘zgaruvchanlik kontseptsiyasi — ayniqsa turlar kabi toifalar ichida ota-onadan naslga o'tadigan tug'ma farqlarning mavjudligi — 18 va 19-asrlarda yashagan kishilar uchun mavjud bo'lmagan zamonaviy genetika g'oyalariga tayanmaydi.
Irsiy oʻzgaruvchanlikning Darvingacha boʻlgan tushunchalari
1700-yillarning oʻrtalarida fransuz olimi Pierre Louis Maupertuis, turlarning haqiqiy, asl shakli boʻlsa-da, rivojlanayotgan nasl davridagi tasodiflar vaqt oʻtishi bilan toʻplanib boradigan oʻzgaruvchanlikni keltirib chiqarishi mumkinligini taxmin qildi. Darhaqiqat, baʼzi tarixchilar uning gʻoyalari Gregor Mendel tomonidan ishlab chiqilgan irsiyat qonunlarini oldindan koʻra olganini taʼkidlaydilar.
Shu bilan birga, fransuz faylasufi Denis Didro irsiy oʻzgaruvchanlikni yaratish uchun boshqacha asosni taklif qildi. Didro Mopertyuining koʻpayish va naslning keyingi oʻsishi paytida oʻzgaruvchanlik paydo boʻlishi mumkinligi haqidagi gʻoyasini oʻzlashtirdi va „normal“ organizm ishlab chiqarish ehtimoli „dahshatli“ (monstrous) organizm ishlab chiqarish ehtimolidan yuqori emas deb hisobladi. Biroq, Didro materiyaning oʻzi hayotga oʻxshash xususiyatlarga ega ekanligiga va hayot potentsialiga ega boʻlgan tuzilmalarga oʻz-oʻzidan yigʻilishi mumkinligiga ishondi Didroning biologik transformatsiya haqidagi gʻoyalari, uning 1749-yildagi "Letter on the Blind" asarida keltirilgan boʻlib, mavjud turlar ichidagi oʻzgaruvchanlikka emas, balki oʻz-oʻzidan paydo boʻlgan shakllarning oʻzgaruvchanligiga qaratilgan edi.
Ham Mopertyui, ham Didro Rim shoiri va faylasufi Lukretsiy gʻoyalariga tayanganlar, u oʻzining „De rerum natura“ asarida butun koinot tasodifiy yaratilganligi va faqat oʻz-oʻziga zid boʻlmagan mavjudotlar omon qolganini yozgan. Mopertyui ishi Lukretsiy va Didro ishlaridan hayot vaqt oʻtishi bilan oʻzgarishiga ishonuvchilar orasida yangi gʻoya — mavjudotlarning tabaqaviy omon qolishini tushuntirishda muvofiqlik (conformity) tushunchasidan foydalanishi bilan ajralib turadi.
Didro singari, 18-asrning boshqa ikki nufuzli aqllari — Erazm Darvin va Jan-Batist Lamark — faqat juda oddiy organizmlar oʻz-oʻzidan paydo boʻlishi mumkinligiga ishonishgan, shuning uchun yerda kuzatilgan murakkab hayotning ulkan xilma-xilligini yaratish uchun boshqa mexanizm kerak edi Erazm Darvin hayvonning hayoti davomida orttirilgan oʻzgarishlar uning nasliga oʻtishi mumkinligini va bu oʻzgarishlar hayvonning asosiy ehtiyojlarini qondirishga boʻlgan intilishlari natijasida paydo boʻlishini taklif qildi. Shunga oʻxshab, Lamarkning tirik mavjudotlar oʻrtasidagi oʻzgaruvchanlik nazariyasi aʼzolardan foydalanish va foydalanmaslik qonuniyatlariga asoslangan boʻlib, u buni irsiy fiziologik oʻzgarishlarga olib keladi deb hisoblagan Ham Erazm Darvin, ham Lamark rivojlanish davrida yoki hayvonning hayoti davomida paydo boʻlgan variatsiya irsiy ekanligiga ishonishgan, bu individlardan populyatsiyalarga oʻtadigan vaqt oʻtishi bilan oʻzgarish nazariyalaridagi muhim qadam edi.
Keyingi asrda Uilyam Gershel teleskop orqali tungi osmondagi turli tumanliklarni kuzatish natijasida har xil tumanliklar kondensatsiya jarayonining turli bosqichlarida boʻlishi mumkinligini taxmin qildi. Nebulyar gipoteza (nebular hypothesis) deb nom olgan ushbu gʻoya tabiiy jarayonlar materiyadan tartib yaratishi va oʻzgaruvchanlikni kiritishi mumkinligini va bu jarayonlarni vaqt oʻtishi bilan kuzatish mumkinligini koʻrsatdi Zamonaviy oʻquvchiga astronomik nazariyalar organik oʻzgaruvchanlik nazariyalariga aloqador emasdek tuyulishi mumkin boʻlsa-da, bu gʻoyalar 19-asr oʻrtalarida biologik transformatsiya — biz hozir biladigan evolyutsiya — gʻoyalari bilan sezilarli darajada birlashib, Charlz Darvin kabi keyingi mutafakkirlar ishi uchun muhim poydevor yaratdi.
Darvinning irsiy o‘zgaruvchanlik tushunchasi
Charlz Darvinning irsiy o‘zgaruvchanlik haqidagi g'oyalari ham uning o'z ilmiy ishlari, ham zamondoshlari va salafiy g'oyalari ta'sirida shakllangan. Darvin irsiy o‘zgaruvchanlikni ko'plab omillar bilan bog'lagan, ammo ayniqsa tanaga ta'sir qiluvchi atrof-muhit kuchlariga urg'u bergan. Uning irsiyat nazariyasi gemmulalar (gemmules) — organizmning mohiyatini qamrab oluvchi va butun tanadan reproduktiv organlarga boradigan va u yerdan naslga o'tadigan gipotetik kichik zarrachalar haqidagi (hozirda rad etilgan) pangenezis (pangenesis) g'oyasiga asoslangan edi. Darvin atrof-muhit va tana o'rtasidagi sabab-oqibat munosabatlari shunchalik murakkabki, bu munosabatlar natijasida hosil bo'lgan variatsiyalar tabiatan oldindan aytib bo'lmaydigan darajada ekanligiga ishongan. Biroq, Lamark singari u ham o'zgaruvchanlik organlardan foydalanish va foydalanmaslik qonuniyatlari natijasida paydo bo'lishi mumkinligini tan olgan. Darvin ham tabiiy, ham xonakilashtirilgan populyatsiyalardagi o'zgaruvchanlikka qiziqqan va populyatsiyadagi individlarning maqsadsiz ko'rinadigan o‘zgaruvchanlikka ega ekanligini anglab etishi ko'p jihatdan uning chorvadorlar bilan ishlash tajribasiga bog'liq bo'lgan. Darvin turlarning kichik va uzluksiz o'zgarishlarning to'planishi orqali asta-sekin o'zgarib borishiga ishongan, bu tushuncha 20-asrgacha qizg'in bahs-munozaralarga sabab bo'lgan.
Irsiy o‘zgaruvchanlikning Darvindan keyingi tushunchalari
20-asrda populyatsion genetika (population genetics) deb nomlanuvchi soha rivojlandi. Ushbu soha genetik o‘zgaruvchanlikni tushunish va miqdoriy jihatdan baholashga intiladi Quyidagi bo'lim populyatsion genetikadagi tanlangan ishlanmalar xronologiyasidan iborat bo'lib, genetik o‘zgaruvchanlikni o'lchash usullariga e'tibor qaratadi.
- 1866 — Geterozigotalilik: Gregor Mendelning duragaylash tajribalari 1950-yillarda geterozigotalilik (heterozygosity) deb e'tirof etilgan kontseptsiyani kiritdiDiploid turda — har bir hujayrasida DNKning ikki nusxasini (har bir ota-onadan bittadan) saqlaydigan turda — agar organizmning DNK nusxalari genomdagi ma'lum bir nuqtada farq qilsa, u individ ushbu saytda geterozigota hisoblanadi. Populyatsiyadagi geterozigotalarning o'rtacha chastotasi bo'lgan geterozigotalilik 20-asr o'rtalariga kelib populyatsiyadagi genetik o‘zgaruvchanlikning asosiy o'lchoviga aylandi... Agar populyatsiyaning geterozigotaliligi nolga teng bo'lsa, har bir individ gomozigotali bo'ladi; ya'ni har bir individ qiziqish uyg'otayotgan lokusda bir xil allelning ikki nusxasiga ega va genetik o‘zgaruvchanlik mavjud emas.
- 1918 — Dispersiya: «Mendel irsiyati faraziga ko'ra qarindoshlar o'rtasidagi korrelyatsiya» deb nomlangan muhim maqolasida R.A. Fisher dispersiya (variance) statistik tushunchasini kiritdi; bu kuzatuvlar to'plamining o'rtacha qiymatdan kvadratik og'ishlarining o'rtacha qiymatidir (), bu yerda — dispersiya va — kuzatuvlari olingan populyatsiyaning o'rtacha qiymatidir. R.A. Fisherning populyatsion genetikadagi ishi nafaqat genetika uchun muhim edi; bu g'oyalar zamonaviy statistikaning asosini ham tashkil etdi.
- 1918, 1921 — Additiv va dominant genetik dispersiya: R.A. Fisher keyinchalik o'zining dispersiya haqidagi umumiy ta'rifini populyatsion genetika uchun tegishli bo'lgan ikki komponentga ajratdi: additiv va dominant genetik dispersiya. Additiv genetik model fenotipga ta'sir qiluvchi genlar soni kichik bo'lsa, genlar bir-biri bilan o'zaro ta'sir qilmaydi va belgi qiymatini har bir genning belgi ustidagi ta'sirini shunchaki yig'ish orqali hisoblash mumkin deb faraz qiladi. Fisher modeli bo'yicha jami genetik dispersiya additiv genetik dispersiya (belgi ustidagi additiv ta'sirlar natijasidagi dispersiya) va dominant genetik dispersiya (genlar o'rtasidagi o'zaro ta'sirlarni hisobga oladi) yig'indisidir
- 1948 — Entropiya: Genetik dispersiyani miqdoriy baholash maqsadida ishlab chiqilgan dispersiyadan farqli o'laroq, Klod Shennonning o‘zgaruvchanlik o'lchovi, hozirda Shennon entropiyasi (Shannon entropy) deb nomlanadi, uning aloqa nazariyasidagi ishining bir qismi sifatida xabarda saqlanadigan ma'lumot miqdorini aniqlash usuli sifatida ishlab chiqilgan. Biroq, bu usul populyatsion genetikada tezda o'z o'rnini topdi va Richard Levontinning «Inson genetik xilma-xilligining taqsimlanishi» nomli muhim maqolasida genetik o‘zgaruvchanlikni aniqlash uchun ishlatilgan asosiy usulga aylandi..
- 1951 — F-statistikalar: F-statistikalar (fixation indices) populyatsion genetik Suoll Rayt tomonidan populyatsiyalar ichidagi va ular orasidagi genetik o‘zgaruvchanlikdagi farqlarni aniqlash uchun ishlab chiqilgan. Ushbu statistikalarning eng keng tarqalgani FST bo'lib, u o'zining eng oddiy ta'rifida bitta genning ikki xil versiyasini yoki allelini hamda ushbu ikki alleldan birini yoki ikkalasini o'z ichiga olgan ikkita populyatsiyani hisobga oladi. FST ushbu ikki populyatsiya o'rtasidagi genetik o'zgaruvchanlikni ikki populyatsiya bo'yicha geterozigotalarning o'rtacha chastotasini ikkala populyatsiya birlashtirilgan taqdirdagi geterozigotalar chastotasiga nisbatan hisoblash orqali aniqlaydi. F-statistikalar dispersiyaning ierarxik tushunchalarini miqdoriy baholash g'oyasini kiritdi va ko'plab muhim populyatsion genetik usullarning, jumladan genomda tabiiy tanlanish belgilarini tekshiradigan usullar to'plamining asosiga aylandi.




Qo'shimcha adabiyotlar
- Mayr E. (1970): Populations, species, and evolution – An abridgment of Animal species and evolution. The Belknap Press of Harvard University Press, Cambridge, Massachusetts and London, England, ISBN 0-674-69013-3.
- Dobzhansky T. (1970): Genetics of the evolutionary process. Columbia, New York, ISBN 0-231-02837-7.
- McGinley, Mark; J. Emmett Duffy (ed). 2008. "Genetic variation." In: Encyclopedia of Earth. Washington, D.C.: National Council for Science and the Environment.
- "Genetic Variation" in Griffiths, A.J.F. Modern Genetic Analysis, Vol 2., p. 7
- "How is Genetic Variation Maintained in Populations" in Sadava, D. et al. Life: The Science of Biology, p. 456
- Nevo, E.; Beiles, A. "Genetic variation in nature". Scholarpedia, 6(7):8821. doi:10.4249/scholarpedia.8821
- Hedrick P. (2011): Genetics of populations. Jones & Bartlett Learning, ISBN 978-0-7637-5737-3.
- Albers, Patrick K.; McVean, Gil (2020-01-17). "Dating genomic variants and shared ancestry in population-scale sequencing data". PLOS Biology 18 (1). doi:10.1371/journal.pbio.3000586. ISSN 1545-7885. PMID 31951611. PMC 6992231. //www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=6992231.
- Rieger R. Michaelis A., Green M. M. (1976): Glossary of genetics and cytogenetics: Classical and molecular. Springer-Verlag, Heidelberg - New York, ISBN 3-540-07668-9; ISBN 0-387-07668-9.
- Griffiths, A. J. F. (1999). An Introduction to genetic analysis. W. H. Freeman, San Francisco, ISBN 0-7167-3520-2.
- Cavalli-Sforza L. L., Bodmer W. F. (1999): The genetics of human populations. Dover, Mineola, New York, ISBN 0-486-40693-8.